







With the use of iMETOS® DECISION SUPPORT SYSTEM you can monitor all relevant climatic conditions and soil parameters, and bring your agricultural work management one step higher.
Combine monitored and forecasted data to get a powerful tool, which will help you optimize your production and produce more grapes with less.
| Weather monitoring | Insect monitoring | Crop Monitoring |
 |  |  |
With the help of climatic and soil monitoring and forecasting you can:
WEATHER MONITORING: Be prepared for frost protection. Set an SMS alarm to notify you, when frost conditions are approaching. Temperatures below – 2 °C can be lethal for your vineyard.
FERTILIZING MANAGEMENT: Optimize fertilization time. The best is if the spreading of the fertilizer is followed by rainfall.
NUTRITION MANAGEMENT: Quick soil analysis helps you apply the appropriate amount of nutrients.
PLANT PROTECTION MANAGEMENT: With the help of disease models, spraying window forecast, rain washout prediction and leaf growth prediction you can optimize your plant protection activities in a way to have disease free grapes.
For example: If spraying is followed by a rainfall of 30 mm, there is a good chance that it will wash away the treatment and you will have to repeat the spraying to keep your grapes protected.
IRRIGATION MANAGEMENT: Optimize irrigation water use by monitoring soil moisture and with use of our water balance model.
HARVEST MANAGEMENT: Monitor air temperature and relative humidity in the nights before grape harvest.
With CropVIEW® camera device you can have a remote view of your vineyard, directly from your computer or phone, and see at any time how your vine grape is growing and developing.
This is just a glance at solutions that iMETOS® DECISION SUPPORT SYSTEM can bring to you and help you to take control of your vineyard.
- DOWNY MILDEW – Plasmopara viticola
- POWDERY MILDEW – Uncinula necator
- GRAPEVINE BLACK ROT – Guignardia bidwellii
- GRAPEVINE BERRY MOTHS – Lobesia botrana
- GREY MOULD – Botrytis cinerea
- ANTHRACNOSE – Elsinoe ampelina
DOWNY MILDEW
TABLE OF CONTENT
• Biology of Plasmopara viticola (Downy Mildew)
• Primary Infection in detail
• Secondary Infections (Downy Mildew)
• Practical work
Biology of Plasmopara viticola (Downy Mildew)
P. viticola is an obligate parasite. Which means that green, fresh vine organs are needed to grow. During the vegetation free period it persists forming fruiting bodies, so called oospores. Oospores of oomycetes can survive very long periods in soil. Therefore, we can find downy mildew in places where infections are not possible in every year. In spring when the top soil is moist and warm enough, the oospores will form so called macrosporangia which can release up to 200 zoospores into free water. The zoospores move up to the leaves and clusters by wind in water droplets. They do have two flagella and they move in a water film on the downside of the leaves or the clusters and young berries to find a stoma to enter plant tissue. They enter and germinate into the stoma, in which they transfer all their plasma within less than one hour. In microscopic studies, the finding of stoma, encystation and the germination into the stoma was finished within 90 minutes.
P. viticola grows in the intercellular space and it feeds itself with haustorias penetrating the epidermic and parenchymal cells. In dependence of temperature and relative humidity it develops enough intercellular growth with enough haustorias to form a substomatel body which fills up the whole substomatel area and which lifts up the epidermal tissue from the parenchymal tissue. This leads to the visible symptom of the oil spot.
Oomycetes are sporulating in the absence of light when relative humidity is very high. In P. viticola there is no sporulation if temperatures are below 12°C and relative humidity is below 95%. Sporangiaphores are formed by the substomatel vesicles and they will come out of the stoma. The fresh formed sporangia are sticky and can only be removed from the sporangiaphores by water. During the decrease of relative humidity, the sporangia become try and could be removed by wind too.
Sporangia will release up to 20 zoospores into free water. These zoospores have to be distributed by wind in water droplets too to come to fresh leaves, or the sporangia can be distributed by rain or wind itself. The infection process of primary and secondary infection is the same.
Do to the big importance of the sexual stage for the hibernation of the pathogen we can assume mating types fitting in all vineyards where grape vine downy mildew occurs. The zoospore formation takes place on older leafs during summer and early autumn.
Primary Infection in detail
Infections coming from the oospores are called primary infections. This term is misleading since several primary infections can occur in early summer. In epidemiological events, the primary infection does not play an important role if there are sufficient oil spots in the vineyards and the infection potential of the summer spores (sporangia) exceeds the oospores.

The overwintering oospores develop to so-called macrosporangia’s whenever there is sufficient relative humidity for about 24 hours. The macrosporangia’s release their zoospores into free water and heavy rain (as in a thunderstorm). Spores are carried to the vine leaves and/or green shoots. Primary infections, therefore, need longer rainy periods or several successive thunderstorms. The first rain causes the fallen leaves to be saturated with water and strong rain on the following days causes the macrosporangia’s to release their zoospores, which reach the vine leaves or shoots. For the successful infection, a sufficiently long leaf wetness period is necessary to allow the zoospores to reach the stoma of the leaves or shoots and infect them (the Illustration above shows the development cycle of the downy mildew of the vine (Plasmopara viticola).
The Model for the Downy Mildew primary infection checks first, if the weather is suitable for development of macrosporangia’s. This is the case as long as the leaves are wet, or the relative humidity after the rain does not fall below 70%. Depending on the temperature, the macrosporangia’s can develop within 16 to 24 hours. If mature macrosporangia’s are available, it will be shown graphically in the display of the Downy Mildew primary infection. If macrosporangia’s are present, a strong rainfall can spread their zoospores. A continuous rain of 5 mm is interpreted as a strong rain and the zoospores are spread, a primary infection is than possible if the leaf wetness has lasted long enough.
Secondary Infections
Secondary infections of Plasmopara viticola are only possible if already mature oil spots exist in or nearby your vineyard. Oil spots are mature when they are able to sporulate (produce sporangia). Sporangia are only produced by night. Sunlight inhibits the sporulation. Sporangia are produced if it is warmer than 12°C and the relative humidity is very high. The rate of sporangia production increases with temperature up to 24°C. The optimal temperature for sporulation on European grape varieties (Vitis vinifera) is about 24°C. If temperatures exceed 29°C, no sporulation can take place. In our model we check if humidity of more than 95% occurs during the night. If this condition lasts for an accumulated hourly temperature of more than 50°C, the sporulation finished and new sporangia of Plasmopara exist in the vineyard. For example, 50°C hours correspond to 4 hours with 13°C or 3 hours with 17°C.
The following graph show the relation between temperature and wet conditions (relative humidity, leaf wetness, precipitation) which are used to model the infection events in ng.fieldclimate.com.


Sporangia of Plasmopara viticola have a limited lifetime. The warmer it is and the drier the air, the faster they die (in ng.fieldclimate.com we reset to 0 when r.h. is below 50%). They definitely die in the next dew or leaf wetness period, which is too short for infection. The fitness of older sporangia is quite limited therefore. Our model assumes the lifetime of sporangia to be limited to one day.
In order to cause new infections, the sporangia must be distributed. There are two ways for distributing spores: If it rains immediately after the forming of sporangia, they spread with the rain splashes. If the vine leaves stay long enough wet, a high level of Plasmopara viticola infections takes place. If the next morning begins without rain and with decreasing humidity, the dried up sporangia abscise from their branches. Even a slight air movement will carry them to healthy leaves. Unless it rains soon, they will die.
Practical work
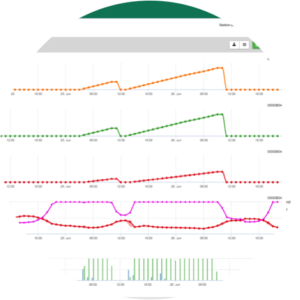
 Graphs in FieldClimate presenting Downy mildew risks
Graphs in FieldClimate presenting Downy mildew risks
Downy Mildew Primary Infection: Infections are shown after macrospore development (orange, fifth graph) in three different severity classes (weak: orange, moderate: green, severe: red) depending on the rain amount and therefore distribution of spores in the vineyard. If 100% infection is determinated optimal conditions have been given to enter the plant tissue (plant protection curative) and the incubation curve (first graph) starts to increase. If the incubation finish (100%) and no plant protection has been taken into account symptoms (oil spots) should have been checked in the field.
1. The infection progress curve and after full infection the consequent incubation period (time between infection and visible symptoms in the field).
2. An infection progress curve is determined when macrosporangia or sporangia are present and leaves are wet. It increases with ongoing leaf wetness and temperature. If the infection progress curve reaches 100% the infection can be assumed to be completed. The incubation progress curve which belongs to the specific infection is started to be calculated with the start of an infection. If an infection can not be completed, the incubation curve will be stopped too.
3. The weather conditions are displayed on the same page with the disease model results. Therefore, you can check with one glance for temperature, relative humidity, rain and leaf wetness during the infection.
4. The model is pointing out infection dates for primary and secondary grape vine downy mildew infections. This is the most critical information. Infections, which have not yet been covered by either preventative or curative fungicides, can be disastrous for the yield/ the quality. Further on information over infection severities (weak/moderate/severe) could be helpful to decide plant protection strategy in dependence of the history of vineyard (amount of inoculum), variety and experience the years before.
Plant protection strategies:
- Prophylactic and systemic or curative fungicides are widely used for the control of downy mildew. Prophylactic chemicals are applied before, but as close as possible to an infection event in the period of greatest host susceptibility, between shoot length of 10 cm and pea-sized berries.
- Prophylactic chemicals maybe used also by spraying as close as possible before a forecast weather event of more than 2mm rain (for primary circle).
- Curative fungicide should be applied as soon as possible after an infection event (100% infection) and before oilspots appear. Be aware of preventing the development of fungicide resistance.
- Monitoring of the vineyard for the presence of downy mildew (oilspots) should be done regularly and taking at least 200 vines into account. A risk is considered, if more than two oilspots per 50 vines are seen.
The model bases on the publications of MUELLER-THURGAU, ARENS, MUELLER and SLEUMER, BLAESER, HERZOG, GEHMANN and many other workers connected to research network on grape vine downy and powdery mildew epidemiology since the early nineteenth of the last century.
POWDERY MILDEW
TABLE OF CONTENT
• Biology of Uncinula necator
• Practical work
• California risk model (by Thomas Gubler)
• Pessl instruments risk model
• Third risk model: Primary inoculum of ascospored
• Practical use
Biology of Uncinula necator
Grape Vine Powdery mildew is caused by the fungus Uncinula necator. It is an obligate parasite, which means it needs green host tissue to feed. U. necator entered Europe in the nineteenth century and it spread through all wine growing countries up to the end of this century. It has very strong effects on yield and it can lead to defoliation of the vines if there is no chemical control. All European grape vine varieties are susceptible to this pathogen. The breeding on resistance against this pathogen is quite new and the results will take more years of work.
U. necator has to overcome the vegetation free period in the most vine growing areas. To do that the pathogen use two mechanism: a) surviving in its fruiting bodies, the so called cleistothecias or b) survive winters too encapsulated in dormant buds where it finds living tissue during the whole winter. The presence of only one or of both modes of hibernation within one vineyard is based in the disease history of this vineyard. Buds can only be infected as long as they are not closed and surrounded by a ring of senescent tissue. This is the case for only several weeks after the formation. In ordinary vine growing situation vines are pruned back to branches with less than 14 buds or to spurs. These remaining buds can only be infected until they bloom. On the other side we can find flag shoots from stage BBCH 15 to 19. Flag shoots are partly or complete mildew covered shoots formed from latent infected buds. These flag shoots are an early and very potent inoculum source. They are mainly not distributed inside a vineyard. This leads to spots with early and high disease incidence inside vineyards and once with low or moderate disease early in the season. Flag shoots are representing only one mating type due to their vegetative history. This can lead to vineyards where we find only few or no cleistothecias due to the dominance of one single mating type.
Where cleistothecia is the only inoculum we will find an ongoing history in cleistothecia hibernation which can only change to flag shoots in years where the disease builds up very early. Cleistothecia formation is good visible due to the yellow coloring of the sexual combined tissue. It can be found in unsprayed vineyards from mid-summer on. In sprayed vineyards it needs longer to build up the disease incidence allowing the mating types to meet. The spread out of Uncinula necator during the summer is influenced primarily by temperature.
The optimal temperatures for the Powdery Mildew are between 21°C and 32°C. The greater the number of hours daily within this temperature range, the higher the risk for Powdery Mildew. Conidia formation, infection and speed of incubation need similar temperature ranges. Due to the fact that U. necator needs no free water for infection and no high relative humidity for conidia formation there is no useable impact of moisture parameter direct on the pathogen.
The presence of free moisture interacts with U. necator through its common hyperparasite Ampelomyces quisqualis. This fungal hyperparasite growth inside the hyphae of powdery mildew fungus. It transforms conidia and cleistothecia into pycnidia of its own. It needs free moisture or very high relative humidity to infect and to grow with thin fast growing hyphae on leaves to find powdery mildew lesions. Heavily infected U. necator lesions are turning the color into a very dark brown. During rain period, they are covered by white spore exudate coming from the pycnidia of the hyper parasite. Ampelomyces quisqualis is available as a commercial product for powdery mildew control too.
Practical work
We have implemented different models regarding infection process into FieldClimate.
a) the fungus overwinters mainly in form of a mycelium. In spring the mycelium starts to grow and built conidiophores with conidia (so no primary infection).
b) if the fungus develops in autumn on berries the sexual form could be present (building cleistothecia) and so in spring the release of ascospores could lead to primary infection (use: model POWDERY MILDEW/ASCOSPORE INFECTION).
California risk model (by Thomas Gubler)
The UC Davis Powdery Mildew Risk Assessment Model (Gubler -Thomas Model) is based on laboratory and field epidemiological studies of grapevine powdery mildew in California.
Risk Assessment Index:
Following ascospore release and germination (use for that model ASCOSPORE INFECTION) , the subsequent development and reproduction of powdery mildew is influenced primarily by ambient temperatures. The UC Davis model evaluates ambient temperatures and assesses the risk of powdery mildew development using a 0-100 point index.
To initiate the risk assessment index, after budbreak and after disease onset, there must be three consecutive days with a minimum of six consecutive hours of temperatures between 21 and 30°C for the powdery mildew epidemic to be initiated. Throughout the calculation of the index, for each day with 6 or more consecutive hours between 21 and 32 °C, the index increases 20 points. The model then fluctuates between 0 and 100 index points based on duration of daily temperatures. The index gains 20 points for each day that meets the requirement of six consecutive hours of temperatures between 21 and 32°C, and it loses 10 points for each day that does not meet the six hour requirement, or if at any time during the day, the temperature accedes over 32°C or is below 21°C.
Interpretation of the index:
Low index values of 0-30 indicate the pathogen is not reproducing. Growers may consider postponing fungicide applications during extended periods with low index values. An index of 40-50 is considered moderate and would imply a powdery mildew reproductive rate of approximately 15 days. Index values of 60-100 indicate that the pathogen is reproducing rapidly (every 5 days) and that the risk for a disease epidemic is great. Graphs in FieldClimate presenting Downy mildew risks
Graphs in FieldClimate presenting Downy mildew risks
Pessl Instruments risk model
The dispersal of Uncinula necator during the summer is influenced primarily by temperature. The antagonist of U. necator called Ampelomyces quisqualis is mainly favored by free moisture. The optimal temperatures for the Powdery Mildew are between 21°C and 32°C. The greater the number of hours daily that the temperature is within this range, the higher is the risk for Powdery Mildew (see model Gubler). Days with more than 6 hours of temperature in this range increase the risk about 20 points. Days where the temperature is not within this optimal range reduces the risk about 10 points. These are days with temperatures under 21°C the whole day long or days where the temperature exceeds 32°C for 6 hours. However days with long leaf wetness periods are leading to an establishment of the antagonistic fungus A. quisqualis. These will lead to an decrease of U. necator.
For this model we recommend following this rules: If the Powdery Mildew risk is less than 20 points spraying interval can be extended. With 20 to 60 points the normal spraying interval is valid. If the risk is more than 60 points you should shorten the spraying interval.
Risk starts: with 3 days with equal or more than 6 hours of: 21°C <= Temperature < 32°C ==> 60 Points
Risk increases: With every day with equal or more than 6 hours of: 21°C <= Temperature < 32°C ==> +20 Points
Risk decreases: With every day when temperature does not reach 21°C or every day with 6 hours of: 32°C <= Temperature and every day where leaf wetness is lasting longer than 8 hours (implementing A. quisqualis) ==> -10 Points

Third risk model: Primary inoculum of ascospores
We recommend to use this model when ascospores are formed during cold winter time (so not infection by mycelium (“Flag shoots”).
Ascospore infection predicts the ascospore release and initial infections, which are based on average temperatures during extended leaf wetness periods.
For calculation in ng.fieldclimate.com approximately 2.5mm of rainfall are required to release ascospores followed by at least 8-12 hours of leaf wetness after ascospore release at temperatures between 10 -15°C for infection. Once infection has occurred, the model switches to a disease risk assessment phase (use model Gubler/Risk model) and is based entirely on the effects of temperature on the reproductive rate of the pathogen. Graphs in FieldClimate presenting Downy mildew risks
Graphs in FieldClimate presenting Downy mildew risks
Practical use
In this model we display the infection by Uncinula necator in a line ranging in between 0 and 100%.
We display two different models in FieldClimate:
1. Possible Ascospore Infections following the models proposed by David Gadoury and Doug Gubler are displayed in the graph for ascospore infection. If the progress curve for the infection model reaches 100% there is the possibility of an ascospore infection. 3 to 5 days after this event first symptoms of powdery mildew might be visible on leaves close to the old cordon.
2. The powdery mildew risk following the California Model and the model for moderate climate zones is displayed in the graph above. Periods with risk have to be covered with fungicide applications. In periods with risk higher than 60 points it is recommended to spray more frequent specially around and short after bloom.
GRAPEVINE BLACK ROT
TABLE OF CONTENT
• Biology
• Practical use
Grape black rot affects many grape growers throughout the United States and most the other warm and arid grape growing areas worldwide. For managing the disease, it is important to understand the disease life cycle and environmental conditions. Once infection takes place, different methods are available to control the disease.
Biology
The grape black rot pathogen overwinters in many parts of the grape vine and is also able to over winter on the ground especially in mummies. In addition, pathogen can overwinter for at “least 2 years within lesions of infected shoots that are retained as canes or spurs.”
Once, rains could release the overwintering spores (ascospores) that form within mummies on the ground and can be dispersed by the wind. Some of the mummies on the ground can have a significant discharge of ascospores that begins about 2 to 3 weeks after bud breaks and will mature 1-2weeks after the start of bloom. A second type of spore (Conidia) can also form within cane lesions or mummies that have remained within the “trellis, and these are dispersed short distances (inches to feet) by splashing rain drops.” Infection occurs when either of the spore types land on green grape tissue and tissue remains wet for a “sufficient length of time, which is dependent on temperature.” The period that these overwintering spores are allowed to cause infection depends on the source. If there is a large source for infection, infection will set in early. In the presence of moisture, these ascospores slowly germinate, taking 36 to 48 hours, but eventually penetrates the young leaves and fruit stems (pedicels). The infections become visible after 8 to 25 days. When the weather is moist, ascospores will be produced and released throughout the entire spring and summer, providing continuous primary infection. (Wilcox, Wayne F. “Black rot Guignardia bidwellii.” Disease Identification Sheet No. 102GFSG-D4. 2003. Cornell. 24 Oct. 2010).
So the black rot fungus requires warm weather for optimal growth; cool weather slows its growth. A period of 2 to 3 days of rain, drizzle, or fog is also required for infection.” (Ries, Stephen M. “IPM : Reports on Plant Diseases : Black Rot of Grape.” Integrated Pest Management at the University of Illinois. Dec. 1999. 24 Oct. 2010 ). Raindrops transfer these spores by moving the spores to different plant parts, especially susceptible young leaves. If water is present, the conidia germinate in 10 to 15 hours and penetrate young tissue. New black rot infections continue into late spring and summer during prolonged periods of warm, rainy weather. The conidia are capable of germinating and causing infection several months after being formed. During August, the pycnidia are transformed into an overwintering stage that, in turn, gives rise to pseudothecia within which the spring spores (ascospores) are produced. Research has shown that ascospores are an important source of primary infections in the spring. In the spring during wet weather, the “pycnidia on infected tissues absorb water and conidia are squeezed out. Conidia are splashed about randomly by rain and can infect any young tissue in less than 12 hours at temperatures between 60-90 degrees. (Ellis, Michael A. “Fact sheet Agricultural and Natural Resources: Grape Black Rot.” Department of Plant Pathology. The University of Ohio State Extension. 2008).
Practical use
For our model the basis is the literature published by Spotts (Spotts. R. A. 1977. Effect of leaf wetness duration and temperature on the infectivity of Guignardia). Through discussions with Daniel Molitor we modified the original Spotts model toward a model separating into light, moderate and severe infections similar to the design of Apple Scab models. (Daniel Molitor: Untersuchungen zur Biologie und Bekämpfung der Schwarzfäule (Guignardia bidwellii) an Weinreben. Dissertation Justus-Liebig-Universität Gießen, 2009).
An infection fulfilling the “Spotts criteria” is rated to be a light infection. Infections fulfilling the “Spotts criteria” by 150% are rated to be moderate and infection fulfilling the “Spotts criteria” by 200% are rated to be severe.
The model shows the infection dates for light, moderate and severe infections. When 100% of an infection has been fulfilled conditions for spore discharge, germination and infection of the plant tissue have been given. The severity of the infection depends on the time period of optimal temperature and wetness period of the fungus. In dependence of the infection pressure the infections should be covered preventative or a curative spray has to be applied short after the infection. In the moderate semi-arid climate of the Mosel or the most Austrian wine growing areas a spray after determination of a light infection would not be recommended.

GRAPEVINE BERRY MOTHS
TABLE OF CONTENT
• Biology and life cycle
• Monitoring and management
• Modelling European Grapevine Berry moth in FieldClimate platform
Lobesia botrana (Totricidae, Olethreutinae) was firstly described from Austria, but originated from Southern Italy and immigrated throughout Europe, North and West Africa, the Middle East, and Eastern Russia. It was more recently introduced into Japan, and in 2008 it was first reported in Chile.
Hosts Grape (Vitis vinifera) and spurge laurel (Daphne gnidium) are preferred hosts, but it has also been reported on blackberry (Rubus fruticosus), gooseberry (Ribes sp.), black and red currant (Ribes nigurm), olive (Olea europaea), cherry (Prunus avium), prune (Prunus domestica), persimmon (Diospyrus kakis), kiwi (Actinidia chinensis), pomegranate (Punica granatum), carnation (Dianthus spp.), and a number of other wild hosts.
Damage In May and June, first-generation larvae web and feed on the flower clusters. Second-generation larvae (July-August) feed on green berries. Young larvae penetrate the berry and hollow them out, leaving the skin and seeds. Third-generation larvae (August-September) cause the greatest damage by webbing and feeding inside berries and within bunches, which become contaminated with excrement. Additionally, feeding damage to berries favors infection by Botrytis and other secondary fungi such as Aspergillus, Alternaria, Rhizopus, Cladosporium, and Penicillium.
Biology and life cycle
The adult moth is approximately 6-8 mm long, with a wingspan of 11-13 mm. The female is slightly larger. Both males and females have similar mosaic-patterned wings. The first pair of wings (forewings) is tan-cream in color, mottled with gray-blue, brown, and black blotches. The second pair of wings is gray with a fringed border. The wings are held in a bell shape over the abdomen when at rest. The female lay their eggs separately. Eggs are elliptical and flat, approximately 0.6-0.8 mm in diameter. These lentil-shaped eggs are visible and iridescent creamy white, turning yellow as the embryo develops and later black when the head of the developing larva is formed. There are five larval stages. The first stage is about 1 mm and the last stage about 12-15 mm in size. Larvae are with with a black head and they turn to yellowish brown and the body takes the color of their nutrition (gut content). The fifth last larval instar spin a grayish silicon cocoon to pupate. The pupa is about 4-9 mm long.
In our climatic zone the European grapevine berry moth has two to three generations per year. Four generations are reported from warmer regions of Spain, Greece, Egypt. The pupal stage overwinters inside the silicon cocoons (diapause) under the bark or in soil cracks or on hidden, protected places. When air temperatures increase above 10°C for about 10 to 12 days Adults hatch. First males are found and after one week the female hatch. So, the first male flight may start as early as bud break and the flight continues for about 4 to 5 weeks. Flying activities are when sun rises and temperatures are about 12°C. During the flight the insects mate and the majority of females just mate once. Egg laying activities start shortly after mating (about one to two days). They deposit the eggs singly on or near the flowers. A female lay about 100 eggs and adult life for 1 to three weeks depending on the climatic conditions.
In spring egg hatching takes 10 days or more, while in summer eggs hatch within 3 to 4 days depending on the temperature conditions. The first-generation larvae web on flowers and feed on them, they may enter the peduncle and cause the bunch to dry up. Larval development needs about 20 to 30 days depending on weather conditions. Then they pupate under the bark or in soil or on the flower and adults emerge again 6 to 14 days after.
The second- and third-flight female moths lay eggs individually directly on shaded berries. Shortly after the larva emerges it enters a berry and hollows it out as it feeds. A single bunch may be infested with several larvae. Webbing, frass, and fungal infection may result in extensive contamination of the bunch.
The lower threshold for development is about 10 °C and the upper developmental thresholds is 30°C. Optimal development conditions are between 26-29°C and 40 to 70% humidity. If day length decreases and it is getting colder the diapause stage get initiated. Larval stages die when temperatures fall below 8°C, but the diapausing pupa can withstand even the cold northern European winters.
The first generation is shorter than the summer generations. Eggs hatch in about 66 degree-days Celsius (DDC). Larvae feeding on flower clusters are reported to develop faster than those feeding on grape berries later in the season, and this influences generation time. Non diapausing pupae require about 130 DDC to develop. Adult females may lay eggs about 61 DDC after emergence.
Estimates of DD for a generation vary considerably in the literature, from 427 DDC to 577 DDC in the first generation to 482 DDC to 577 DDC in later generations.
Monitoring and management
Males are attracted by pheromones, which is used to monitor male flights. Before bud break, place traps with L. botrana lures high in the canopy (iScout pheromone) . Place at least one trap per 30 acres or per vineyard block if smaller. Change lures according to manufacturer’s recommendations. Check traps weekly, recording the number of moths caught and removing trapped moths from the sticky trap bottom. Plot the weekly catches to determine initiation and peak of male flights in each generation. Continue monitoring with traps until the peak of the third flight.
Insecticide applications should be timed for larval emergence, thus monitoring egg laying and determining egg hatch are essential to management of this pest. For the first generation, egg laying should be monitored from the peak until the end of the flight. Search for eggs on the peduncle of 100 clusters, selecting one cluster per vine. Note the stage of the majority of the eggs found. Eggs are white when recently laid, turning yellow and later black when larvae are near emergence. A translucent egg chorion indicates the larva has emerged. After egg hatch, look for webbing of flower parts. Open up the webbing and look for feeding damage and larvae.
Begin monitoring for second- and third-generation eggs on berries one week after the first moths of the respective flight are caught in the traps. Continue monitoring for eggs weekly until one week after peak flight. Inspect 100 bunches, selecting one per vine. Continue monitoring bunches for feeding damage (holes or hollow berries), webbing, and presence of larvae.
In countries where L. botrana is established, control measures are targeted at the second generation. This is due in part to the prolonged emergence of the first generation and because of possible reinfestation from untreated neighboring vineyards. However, treatment of the first generation is recommended if populations are high or if treatments are conducted on an area-wide basis. Insecticides are less effective after bunch closure. Several reduced-risk insecticides are registered for use in grapes to control tortricid larvae. These include insect growth regulators, spinosyns, and Bacillus thuringiensis.
Mating disruption has been studied in Europe for several years. It has proven most effective when grapevine moth populations are low and when applied to large areas of over 10 acres or areawide.
Numerous predators and parasitoids are reported in the European literature. Among the parasitoids are 4 species of tachinid flies and nearly 100 species of parasitic wasp in the ichneumonid, braconid, pteromalid and chalicidoid families. The parasites that are reported to cause the greatest impact are those attacking the overwintering pupa. In Spain these include the pteromalids Dibrachys affinis and D. cavus, which are reported to cause up to 70% pupal mortality, whereas in Italy the ichneumonids Dicaelotus inflexus and Campoplex capitator are the most important.
Detailed literature
- Briere JF, Pracros P. 1998. Comparison of temperature-dependent growth models with the development of Lobesia botrana (Lepidoptera: Tortricidae). Environ. Entomol. 27: 94-101.
- Coscollá Ramón R. 1981. Algunas consideraciones sobre la dinámica poblacional de Lobesia botrana Den. Schiff. en las comarcas vitícolas valencianas. Bol. Serv. Plagas 7: 169-184.
- Coscollá Ramón R. 1998. Polillas del racimo (Lobesia botrana Den. Y Shiff.). In Los parasitos de la vid, estrategias de proteccion razonada. Madrid, Spain. pp. 29-42.
- Armendáriz I, Campillo G, Pérez-Sanz A, Capilla C, Juárez JS, Miranda L. 2007. La polilla del racimo (Lobesia botrana) en la D.O. Arribes, años 2004 a 2006. Bol. San Veg. Plagas 33: 477-489.
- Del Tío R, Martínez JL, Ocete R, Ocete ME. 2001. Study of the relationship between sex pheromone trap catches of Lobesia botrana (Den. & Schiff.) (Lep., Tortricidae) and the accumulation of degree-days in Sherry vineyards (SW of Spain). J. Appl. Ent. 125: 9-14.
- Gabel B, Mocko V. 1986. A functional simulation of European vine moth Lobesia botrana Den. Et Schiff. (Lep., Torticidae) population development. J. Appl. Ent. 101: 121-127.
- Gallardo A, Ocete R, López MA, Maistrello L, Ortega F, Semedo A, Soria FJ. 2009. Forecasting the flight activity of Lobesia botrana (Denis & Schiffermüller) (Lepidoptera, Torticidae) in Southwestern Spain. J. Appl. Entomol. 133: 626-632.
- Louis F, Schmidt-Tiedemann A, Schirra KJ. 2002. Control of Sparganothis pilleriana Schiff. and Lobesia botrana (Den. & Schiff). in German vineyards using sex pheromone-mediated mating disruption. Bull. IOBC/WPRS 25: 1-9.
- Maher N. 2002. Sélection du site de ponte chez Lobesia botrana (Lepidoptera: Tortricidae): influence de l’infromation chimique non-volatile présente sur les fruits de plantes hôtes. Thése N° 968, Université de Bordeaux 2. pp. 204 (PDF).
- Maher N, Thiéry D. 2006. Daphne gnidium, a possible native host plant of the European grapevine moth Lobesia botrana, stimulates its oviposition. Is a host shift relevant? Chemoecol. 16: 135-144
- Masante-Roca I, Anton S, Delbac L, Dufour MC, Gadenne C. 2007. Attraction of the grapevine moth to host and non-host plant parts in the wind tunnel: effects of plant phenology, sex, and mating status. Entomol. Exp. Appl. 122: 239-245.
- Milonas PG, Savopoulou-Soultani M, Stavridis DG. 2001. Day-degree models for predicting the generation time and flight activity of local populations of Lobesia botrana (Den. & Schiff.) (Lep., Tortricidae) in Greece. J. Appl. Ent. 125: 515-518.
- Moreau J, Benrey B, Thiéry. 2006. Grape variety affects larval performance and also female reproductive performance of the European grapevine moth Lobesia botrana (Lepidoptera: Tortricidae). Bull. Entomol. Res. 96: 205-212.
- Sáenz-de-Cabezón F, Maron V, Zalom F, Pérez-Moreno I. 2005. Effects of methoxyfenozide on Lobesia botrana (Den & Schiff) (Lepidoptera: Torticidae) egg, larval and adult stages.
- Thiéry D. 2008. Les Tordeuses nuisibles à la vigne. In Les ravageurs de la vigne. Féret, Bordeaux. pp. 15.
- Torres-Vila LM, Stockel J, Roehrich R, Rodríguez-Molina MC. 1997. The relation between dispersal and survival of Lobesia botrana larvae and their density in vine inflorescences. Entomol. Exp. Appl. 84: 109-114.
- Xuéreb A, Thiéry D. 2006. Does natural larval parasitism of Lobesia botrana (Lepidoptera: Torticidae) vary between years, generation, density of the host and vine cultrivar? Bull. Entomol. Res. 96:105-110.
- Zangheri S, Briolini G, Cravedi P, Duso C, Molinari F, Pasqualini E. 1992. Lobesia botrana (Denis & Schiffermüller). In Lepidotteri dei fruttiferi e della vite. Milan, Italy. Pp. 85-88.
Modelling European grapevine berry moth in FieldClimate platform
Sensor needed: Air temperature
Hourly data Output: flight activity, egg laying activity, first generation, second generation and third generation, accumulated degree days
For flight activity, egg and good egg laying activities: we check the data of the last 8 hours (between 19:00 p.m. and 03:00 a.m.) Threshold for flight activity is >12°C, for egg laying activity > 15°C and for good egg laying activity > 17°C. If all conditions for egg laying activity have been fulfilled we generate the first generation (= accumulated degree days). First generation +530 DD is second generation (if not predicted because of conditions before). Second generation + 530 DD is third generation.
In the graph you see the occurrence of the first generation (good conditions for all three activities of flight, egg laying and good egg laying) on about 469 degree-days accumulated. The flying of the second generation starts on the middle of June at about 1010-degree days accumulated and good temperature conditions for egg laying. Graphs in FieldClimate presenting Downy mildew risks
Graphs in FieldClimate presenting Downy mildew risks
GREY MOULD
TABLE OF CONTENT
• Biology of B. cinerea
• Model of B. cinerea and practical use
Botrytis cinerea is a necrotrophic fungus that affects many plant species, although its most notable hosts may be grapes.
In viticulture, it is commonly known as botrytis bunch rot; in horticulture, it is usually called grey mould or gray mold.
The fungus gives rise to two different kinds of infections on grapes. The first, grey rot, is the result of consistently wet or humid conditions, and typically results in the loss of the affected bunches. The second, noble rot, occurs when drier conditions follow wetter, and can result in distinctive sweet dessert wines, such as Sauternes or the Aszú of Tokaj. The species name Botrytis cinerea is derived from the Latin for “grapes like ashes”; although poetic, the “grapes” refers to the bunching of the fungal spores on their conidiophores, and “ashes” just refers to the greyish color of the spore masses. The fungus is usually referred to by its anamorph (asexual form) name, because the sexual phase is rarely observed. The teleomorph (sexual form) is an ascomycete, Botryotinia cinerea.
Biology of B. cinerea
In fruits and grapes flower infections followed by latency are having a big impact onto the epidemiology of grey mould. Several flower to fruit infection pathways can be separated. In grapes, kiwifruits, raspberries infections trough the stylus into the ovule have been postulated. In the ovulus the pathogen stays latent, which seems to be a result of preformatted host defense strategy (similar to the resveratrol content of young grape berry). In grapes, kiwifruits and strawberries infections trough the stamen, petals or septals have been found important. In grapes studies showed that B. cinerea could infect the stamens and grows basipetally to infect the receptacle and then grows systemically to the pedicel and vascular tissues in the berries.
A 6-year research project in the Cape area showed that grape berries can be infected through the stoma and lenticels of the pedicel and at a lower extend of the rachis. Pedicel infections are possible during flowering period too. Later this tissue increases resistance against B. cinerea infections.
Other infection pathways postulating the saprophytic growth of the pathogen on floral debris and the later infection of berries when the susceptibility increases with ripening or by insect or hail damage of the berries. Assumption of conidia accumulation within the fruit during summer and the infection of susceptible berries later in the season is another thesis. Conidial infection of ripening fruit is possible from any inoculum source. Most probably a low number of latent infected berries are formed, which show extensive sporulation when susceptibility of the berries increases with maturity. With can assume berries to become susceptible starting from a sugar content of 6%.
In kiwi fruit we have a big impact of picking conditions to the occurrence of B. cinerea. Fruits picked with a wet surface can get infected by B. cinerea at the micro lesions set by the fingers of the pickers.
In practical control of B. cinerea we have to separate two important infection periods: Flowering and senescence. 1) During flowering we have susceptible young fruits, where the infection is followed by a latency period. 2) While infection on matures (senescent) fruits will lead to symptoms without latency period. The importance of the infection during flowering in grapes can change from season to season and between region. In fruits where we have to expect some shelf live (table grapes, kiwis or strawberries) the symptoms are seen when stored under cooled conditions in shops or storages. A chemical control of vine grapes showing a good resistance to B. cinerea during the flowering will not show any economic results. Therefore, all conditions of risk and probability of infection, the susceptibility of the fruit and the shelf life, storage conditions have to be taken into consideration in the decision of an application against Botrytis cinerea during flowering.
In stone fruits infection by B. cinerea occurs mostly during flowering. At this time treatments against Monilina spp. are taken into considerations which also infect the Botrytis cinerea infection.
Model of B. cinerea and practical use
Sensors needed: Leaf wetness, temperature, relative humidity
Botrytis cinerea is a facultative parasite. It grows on dead plant material too. Because of this fact it is always present in vineyards and orchards. Botrytis cinerea is related to moist climate. For infection it needs very high relative humidity or the presence of free water (sensor: leaf wetness, relative humidity). The fungus is unable to infect healthy adult plant material by spores. Infection takes place on young shoots of the vine during longer wet periods or an shoots damaged by hail storms.
The model use the following correlation between leaf wetness duration and temperature to calculate the risk of an infection.
Graphs in FieldClimate presenting Downy mildew risks
The Botrytis Risk Model results in a risk value of 0 to 100%. This value indicates the pressure of B. cinerea at the time. If we have a value of 100% it means that there has been several times a wetness period long enough to infect the susceptible tissue (we calculate so called “wet points” (array between leaf wetness, temperature with a maximum of initially 38400 points (beginning of season, which displays 30% risk). After this period each wet period with about 4000 wet points (array) increase the risk with 10% or on the other side each dry period reduce the risk by 1/5 of the former value.
An application against B. cinerea is depending on the fruit and the production target.

ANTHRACNOSE
TABLE OF CONTENT
• Symptoms
• Life cycle of the pathogen
• Modelling in Fieldclimate.com
Anthracnose, black spot, bird`s eye rot, bird`s eye spot is found worldwide. The disease reduces fruit quality and yield, weaken the vine via significant destruction of new shoots and leaves. Most Vitis species are susceptible to anthracnose.
In humid grape vine growing seasons the disease is well established, it causes damages to highly susceptible cultivars following early season rains.
Once established in vineyards, the disease can be very difficult to manage. However, in Europe the disease is nearly negligible because of fungicides with copper compounds, dithiocarbamates, phthalimids to control downy mildew and the lack of rainfall in the arid climates.
Symptoms
The disease attacks the aerial, succulent parts of the vine, including young shoots, leaves, petioles, tendrils and clusters; lesions on shoots and berries are most common.
First symptoms appear as isolated, small, circular spots which become brownish, sunken with gray centers and dark, round or angular margins. Sometimes symptoms look like hail injury. On leaves numerous small, brown spots appear, which are gray in the center, the necrotic center usually drops out, creating a “shot-hole” appearance.
Young leaves are more susceptible for infections.
On berries small reddish- brown, circular spots develop, which become slightly sunken. The center turns grey and will be surrounded by reddish- brown to black margins (bird`s eye).
Life cycle of the pathogen
The disease is caused by the pathogen Elsinöe ampelina (de Bary) Shear.
Infected canes are the main source of the disease. Sclerotia or mycelium surviving in the lesions become active in spring and produce conidia under wet conditions (rain or dew for 24 hours) within temperature regime of 2°C-40°C. The conidia are splashed by rain to new tissue. They germinate and produce new infections. Warm weather reduces the wetting time needed for initial infection and the incubation period before symptoms are visible on the leaves. Optimum spore germination occurs at 25-30°C, with a minimum of 3- 4 hours of required leaf wetness. Also, the incubation time is shortest within this temperature range (3-4 days under ideally wet conditions). At infections at temperature around 10°C, disease symptoms need about 14 days to be visible on leaves.
Conidia or ascospores formed on infected berries that overwinter on the vine or floor may also cause primary infections. Spores infect new leaves, shoots, tendrils, young berries and produce lesions at humid conditions. These conidia serve as secondary inoculum and are responsible for further infections in the season. The conidia are dislodged by raindrops and dispersed in the vineyard.
Epidemic development is caused by susceptibility of the grapevine tissue, dispersal of conidia, and prevailing weather conditions. The most important factor is the frequency of rainy periods.
Modelling in Fieldclimate.com
Overwintering spores development: Temperature: 2-40C°, relative humidity above 90% or leaf wetness- when spores are developed (100%) and still conditions of leaf wetness and temperature infection starts to be calculated. Spore development (r.h. below 50% ) and infection stops to be calculated when humidity is too low. Severity of infection depends on the wet conditions (rain event). Graphs in FieldClimate presenting Downy mildew risks
Graphs in FieldClimate presenting Downy mildew risks




With the use of iMETOS® DECISION SUPPORT SYSTEM you can monitor all relevant climatic conditions and soil parameters, and bring your agricultural work management one step higher.
Combine monitored and forecasted data to get a powerful tool, which will help you optimize your production and produce more blueberries with less.
With the help of climatic and soil monitoring and forecasting you can:
- WEATHER MONITORING: Be prepared for frost protection. Set an SMS alarm to notify you, when frost conditions are approaching. Temperatures below – 0,6 °C can be lethal for your blueberry field.
- FERTILIZING MANAGEMENT: Optimize fertilization time. The best is if the spreading of the fertilizer is followed by rainfall.
- PLANT PROTECTION MANAGEMENT: With the help of disease models for blueberries (for anthracnose) and spraying window forecast, you can optimize your plant protection activities in a way to have disease free blueberry yield.
For example: If spraying is followed by a rainfall of 30 mm, there is a good chance that it will wash away the treatment and you will have to repeat the spraying to keep your crop protected. - IRRIGATION MANAGEMENT: Optimize irrigation water use by monitoring soil moisture and with use of our water balance model.
- HARVEST MANAGEMENT: Monitor your soil moisture to know when your crop is suffering water stress and start your irrigation system on time. Also monitor your irrigation water pH to prevent nutrient imbalances.
The fungal pathogen Colletotrichum acutatum causes Blueberry Anthracnose (Ripe Rot) on pre and post-harvest fruit of highbush blueberry. The disease occurs in all blueberry production areas and has a great impact of reducing shelf life of fruits and having a poor quality of fresh fruits on the market.
In FieldClimate weak, moderate and severe Infection is calculated for blueberry anthracnose in dependence of next parameters: temperature, leaf wetness and precipitation. Whenever 100 % of infection is reached, the conditions for a weak, moderate or severe infection have been fulfilled. You can see on graph that the conditions for infection were fulfilled, when the relative humidity was high and leaf wetness was present for a longer period of time.
Click on the graph for a larger picture.


With the CropVIEW® camera device you can have a remote view of your blueberry field, directly from your computer or phone, and see at any time how your blueberries are growing and developing.
This is just a glance at solutions that iMETOS® DECISION SUPPORT SYSTEM can bring to you and help you to take control of your crop.
| Weather monitoring | Insect monitoring | Crop Monitoring |
| | |
BLUEBERRY DISEASE MODELS
BLUEBERRY ANTHRACNOSE
TABLE OF CONTENT
• Symptoms
• Disease cycle
• LiteratureThe fungal pathogen Colletotrichum acutatum causes Blueberry Anthracnose (Ripe Rot) on pre and post-harvest fruit of highbush blueberry. The disease occurs in all blueberry production areas and has a great impact on reducing the shelf life of fruits and having a poor quality of fresh fruits on the market.
Symptoms
Blueberrys are susceptible to the fungal pathogen. The berries don`t develop symptoms until they are mature or ripe. The first symptoms are seen near the end of the blossom on ripe fruits as dark, sunken areas. These infected area may shrivel with time. Within a few days bright salomon- orange coloured, wet spore masses are seen on these sunken areas. The sticky spore masses spread to other berries by rain, irrigation, splashing water, wind and contacts during harvest. The fungal pathogen infects all stages of the plant: leaves, buds, twigs and canes. New shoots and leaves may become blighted in the spring. Twigs are becoming infected by transferring spores from infected buds. Dark brown, canker- like lesions with fruit bodies develop on young canes.
Disease cycle
The fungal pathogen overwinters in twigs and flower buds, spurs and trusses. In spring the fungus produce spores, which are dispersed by rain, irrigation and water- splashes. Infections occur at a temperature optimum of 12- 27°C and a leaf wetness period of more than 12 hours. The fruit can be infected at any time, although infections are just visible (symptoms seen) when the fruit is mature. The fungus enters the fruit and remains dormant until the berry starts to ripen. At this time the fungus starts to colonize the infected area and produce enzymes with destroy the plant cell walls. These damaged cells loose their contents and integrity (symptoms are seen as sunken areas and shrivelled). Spent fruit trusses could be infected after harvest. In FieldClimate weak, moderate and severe Infection are calculated in dependance of the parameters temperature, leaf wetness and precipitation. Whenever 100 % of infection are reached the conditions for an weak/moderate or severe infection have been fullfilled.
Literature
Peres N.A., Timmer L.W., Adaskaveg J.E., Correll J.C. (2005): Lifestyles of Colletotrichum acutatum. Plant Disease, Vol.89 (8), 784-796.
Wharton P.S., Dièguez-Uribeondo(2004): The biology of Colletotricum acutatum. Anales del Jardin Botanicode Madrid 61(1).
Gillett J.M., Schilder A. C. (2009): Environmental Requirements for Infection of Blueberry Fruit by Colletotricum acutatum. Proc. IxthIS on Vaccinium.
